
- 发布日期:2024-10-07 06:04 点击次数:200

拍浮领路是鱼类集群、快速追击捕食或规避敌害、生殖洄游、穿越激流与堤坝等生态流程结束的主要神气[1—2], 为其生涯、滋长及衍生提供了基本保证, 具有雄壮的生态学真谛。因此, 拍浮才智也被看作是表征鱼类生涯稳妥度的要道组分。感应流速(Inductive velocity)、快速启动拍浮(Fast-start swimming)、临界拍浮速率(Critical swimming speed, Ucrit)和爆发拍浮速率(Burst swimming speed, Uburst)等是评估鱼类拍浮性能的几个要道计划[3]。平庸觉得鱼类的长距离生殖与索饵洄游才智主要与有氧领路才智(举例Ucrit)关联; 而鱼类完成拿获猎物与走避猎食者、穿越激流寻觅合乎生境等主要与无氧领路才智(举例Uburst)关联[4—5]。水利水电开导等涉水工程建设继续形成山区溪流连通性裁汰校园春色 亚洲色图, 导致鱼类生境片断化、落空化等一系列负面生态问题[6—8]。栖息地破碎对溪流鱼类(尤其是洄游性鱼类)的生涯衍生组成了严重要挟, 同期也对鱼类爆发拍浮才智提议了更高条款, 关联溪流生态系统要道鱼类爆发拍浮才智过头种内种间互异的相关是开展种群保护、栖息地惩处与生态成立的基础与前提[9—10]。
秦岭细鳞鲑(Brachymystax tsinlingensis)从属于鲑形目(Salmoniformes)、鲑科(Salmonidae)、细鳞鲑属(Brachymystax), 俗称梅花鱼, 为我国稀薄迥殊种、国度Ⅱ级重心保护野纯真物。该物种是第四纪冰川残留种、北半球鲑科鱼类中漫步在最南端的两物种之一, 为典型的陆封型冷水性山麓鱼类, 主要漫步在秦岭海拔1000 m以上的山涧溪流中, 漫步区触及长江流域汉江水系(湑水河、酉水河、子午河等)以及黄河流域渭河水系(石头河、千河、黑河等)[11—16]。连年来, 由于环球景象变化以及东谈主类活动按捺形成生涯环境恶化, 秦岭细鳞鲑的漫步范围呈现碎屑化趋势、野生种群数目暴减[12]。打听相关发现, 秦岭细鳞鲑生境特殊, 水文地舆环境复杂, 多大高差地貌与湍急激流, 常需要进行屡次重复爆发拍浮领路以穿越激流甚而跳跃堤坝防碍, 因此爆发拍浮才智过头可握续性对该物种生涯至关雄壮。然而, 对该物种的相关聚集于资源打听、物种武断、驯养生息、种群遗传和热生物学等方面[13—14], 对其不同生活史阶段该拍浮性能(尤其爆发拍浮才智)于今尚无干系相关。
秦岭细鳞鲑栖息地鱼类群落结构苟简, 同域漫步鱼类主要为拉氏鱥(Phoxinus lagowskii)和少许鳅类。拉氏鱥从属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鱥属(Phoxinus), 为袖珍野生经济鱼类, 杂食性, 主要以水生虫豸、软体动物、浮游动物及水生植物为食; 漫步范围较广, 常成群生活于水流较急、清爽、溶氧量高、温度低的山区河沟与小溪环境[17—18]。秦岭细鳞鲑发育较慢, 平庸三龄性熟练, 而拉氏鱥袖珍成体、二龄即可性熟练, 二者在生境选定上具有一定的相同性, 但其栖息地重迭进程依生活史阶段不同而有较大互异[18—19]。一般觉得, 秦岭细鳞鲑处于溪流生态系统食品链尖端、是顶级捕食者, 拉氏鱥是秦岭细鳞鲑主要的饵料生物之一。然而, 看成同域漫步物种, 二者的种间关系又在很猛进程上受到生活史动态变化和栖息地重迭进程的影响; 相等是, 尽管拉氏鱥处于被捕食者地位, 但其可能对秦岭细鳞鲑胚胎形成捕食压力、且对早期生活史阶段秦岭细鳞鲑幼体在食品质源和生涯空间等多方面组成竞争。鉴于此, 揭示两物种不同生活史阶段拍浮才智的动态变化过头种间互异就成为雄壮值得探究的科知识题。在种间水平上, 看成捕食关系详细、栖息环境与生涯需求相异的同域物种, 环境选定压力是否驱动二者进化出不同的爆发拍浮性能?在种内水平上, 不同生活史阶段秦岭细鳞鲑生态习性不同、栖息环境存在较大互异, 而各生活史阶段拉氏鱥栖息环境相对趋于交叠, 生活史阶段效应是否驱动两物种爆发拍浮性能弘扬出不同的种内变异方法?
为探究秦岭细鳞鲑过头同域物种拉氏鱥爆发拍浮才智的种内种间互异, 本相关禁受平允的仿生态鱼类爆发拍浮才智测定安设, 分别测定了不同生活史阶段两种践诺鱼的相对和完全Uburst过头可握续性, 以期为秦岭溪流生态系统生物种种性保护以及将来秦岭细鳞鲑栖息地生态成立提供表面基础。
1 材料与步伐 1.1 践诺动物践诺鱼收集自陕西陇县秦岭细鳞鲑国度级当然保护区科研基地, 共收集到健康轩敞的秦岭细鳞鲑63尾、拉氏鱥64尾, 二者皆漫步于蒲峪河流域(渭河水系千河一级支流)。证实两种践诺鱼个体大小过头体长特征[18—19], 将收集到的践诺鱼辞别红不同的生活史阶段, 具体规格见表 1。将践诺鱼依据种类过头生活史阶段分列成不同的践诺组, 分别置于圆形自净化轮回控温水槽(直径2 m, 水深1 m)中驯养适合48 h。驯养用水为践诺鱼当然栖息地经过千里淀净化处理后的河水, 践诺技艺及时监测水槽内水温与溶氧的变化, 将水温规章在(17±0.5)℃, 水体溶氧>7 mg/L, 禁受当然光照。
1.2 爆发拍浮速率测定安设践诺所用的鱼类仿生态拍浮才智测定安设由本相关团队自行研发。该安设主体为一长直型泳谈(图 1), 并专设有适合区、粉饰区和稠密区, 安设底部模拟当然河谈铺设有碎石。泳谈起原处(粉饰区1)上方加盖有遮光板, 并配备可高下拉伸的活动式闸门以及强光刺激安设; 稠密区规格为5 m×0.25 m×1 m; 泳谈很是另成立遮光粉饰区(粉饰区2)。泳谈旁平行成立5 m钢直尺, 以及时监测践诺鱼位移变动; 稠密区上方架设有与电脑聚拢的摄像头(Logitech Webcam Pro 920), 摄像头距离水面2 m。安设中水深10 cm, 由轮回控温系统将水温规章在(17±0.5)℃, 水体溶氧>7 mg/L, 泳谈末端提供微水流(流速约3 cm/s)。
1.3 爆发拍浮速率测定决策将单尾践诺鱼置入鱼类仿生态拍浮才智测定安设的适合区(图 1)并使其适合15 min, 之后运行爆发拍浮速率测定。测定时, 快速揭开粉饰区1上方的遮光板, 开启强光刺激安设并用玻璃棒对践诺鱼进行追捕刺激, 引导践诺鱼产生爆发拍浮。测按技艺全程摄像, 为考据践诺鱼爆发拍浮速率测定的可握续性, 对每尾践诺鱼连气儿两次进行测定, 两次测定的时辰远离为10 min。
禁受视频处理软件Adobe Premiere Pro对录制视频进行领会。践诺录制视频为30帧/s, 按帧数播放并定位每帧画面中践诺鱼的位置, 以每3帧(即0.1 s)为单元索取践诺鱼爆发领路的距离参数, 选取距离最长的一段用于操办其完全和相对爆发拍浮速率。
完全爆发拍浮速率(Uburst)操办公式如下:
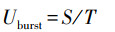
式中: Uburst为完全爆发拍浮速率(cm/s), S是践诺鱼瞬时爆发拍浮冲刺的距离(cm), T是践诺鱼爆发拍浮逃跑的时辰(s)。
狠撸撸相对爆发拍浮速率(rUburst)操办公式如下:
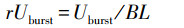
式中, rUburst(Relative burst swimming speed)为相对爆发拍浮速率(BL/s), Uburst为完全爆发拍浮速率(cm/s), BL为践诺鱼体长(cm)。
1.4 数据处理禁受SPSS软件(SPSS 20.0 Inc., USA)对数据进行统计分析。领先测验数据是否降服正态漫步(Shapiro-Wilks测验)及方差皆性。通过操办组内干系统统(Intraclass correlation coefficient, ICC)评估践诺鱼爆发拍浮速率(Uburst和rUburst)的可握续性; 禁受双因素方差分析(Two-way ANOVA)测验物种和生活史阶段对Uburst和rUburst的影响, 禁受单因素方差分析(One-way ANOVA)和Tukey′s HSD多重比较测验物种间以及种内不同生活史阶段的互异。数据均以平均值±圭臬误(Mean±SE)示意, 显贵性水平设为P < 0.05。
2 末端与分析 2.1 爆发拍浮才智测定的可握续性重复测定对不同生活史阶段秦岭细鳞鲑的Uburst和rUburst均无显贵影响(P>0.05, 表 2, 图 2);重复测定对不同生活史阶段拉氏鱥的Uburst和rUburst均影响显贵, 导致其Uburst和rUburst均显贵低于初度测定(P < 0.05, 表 2, 图 3)。
对不同生活史阶段两种践诺鱼Uburst两次连气儿测定组内干系统统ICC的分析末端见表 3。经测验, 统统参数两次连气儿测定值均显贵干系(P < 0.001)且ICC统统均大于0.75, 标明不同生活史阶段两种践诺鱼拍浮才智均具有可握续性(图 2, 图 3)。
2.2 物种和生活史阶段对爆发拍浮才智的影响物种、生活史阶段过头交互作用对践诺鱼Uburst和rUburst均影响显贵(P < 0.05, 表 4)。
关于Uburst, 不管初度仍是重复测定, 在疏通的生活史阶段种间Uburst均互异显贵(P < 0.05, 表 5), 弘扬为: 秦岭细鳞鲑>拉氏鱥(图 4);种内不同生活史阶段Uburst均互异显贵(P < 0.05, 表 6), 均弘扬为: 成体>亚成体>幼体(图 4)。
关于rUburst, 不管初度仍是重复测定, 成体rUburst种间均无显贵互异(P>0.05, 表 5), 初度测定亚成体rUburst种间互异显贵(P < 0.05)而重复测定后种间无显贵互异(P>0.05, 表 5);种内不同生活史阶段rUburst均互异显贵(P < 0.05, 表 6), 弘扬为: 成体 < 亚成体 < 幼体(图 5)。
3 计议 3.1 Uburst测定步伐的可靠性与可重复性当今, 对鱼类拍浮才智的测定尚无长入的圭臬化践诺安设。对Ucrit测定国外上平庸禁受“流速递加法”[20], 但由于爆发拍浮领路的非褂讪特质, 禁受该步伐估量鱼类爆发拍浮才智难度较大[21], 具体测定中相关者巨额禁受匀加快拍浮速率(Constant acceleration test speed, Ucat)示意或替代践诺鱼的Uburst[22—23]。Ucat测定步伐为将践诺鱼置入拍浮管、适合后以某一恒定的加快度握续加多水流速率直至鱼体到达力竭景象, 该步伐的优点是测试环境褂讪、测试末端具有爽朗的可重复性, 其弊端是Ucat完全依赖于握续的水流刺激引导、阑珊有余的生态关联。本相关发明了一种仿生态鱼类爆发拍浮才智测定安设, 该安设最猛进程地模拟践诺鱼田野生境, 通过突发性的刺激引导践诺鱼自主完成爆发逃跑流程。本相关发现, 不同组别鱼类两次测定末端的ICC统统均大于0.75(表 3), 标明该测定步伐具有较好的可靠性与可重复性[24]。
重复拍浮弘扬可用于评估表现于应激源后鱼类生感性能的规复率, 被觉得是比单一领路测试更明锐的生态功能计划[25]。Killen等[26]相关标明, 饱足摄食组欧洲鲈(Dicentrarchus labrax)重复拍浮弘扬显着高于饥饿组和低摄食水平组; Wagner等[27]发现转变饲料脂肪酸组成会裁汰大欧好意思鲑(Salmo salar)在重复测定中的拍浮弘扬, 臆测这可能与食品脂肪身分影响能量产生关联。本相关中, 不同生活史阶段秦岭细鳞鲑Uburst和rUburst的初度测定和重复测定末端均无显贵互异, 而拉氏鱥成体和亚成体重复测定末端均显贵低于初度测定(表 2), 标明与拉氏鱥比较、秦岭细鳞鲑无氧代谢规复才智更强, 臆测Uburst规复力的种间互异可能与两物种异化的生境选定与环境适合神气关联(田野生境下秦岭细鳞鲑需要反复地穿越激流甚而进取跃起穿过叠水区寻找产卵场或其它合乎生境, 而拉氏鱥对此并无横暴生态需求); 联紧缚尾与彭明锐等[28]对秦岭细鳞鲑步法退换速率(Gait transition speed)以及Ucat的测定末端左近。
3.2 两种践诺鱼Uburst的种间互异当然选定压力塑造鱼类拍浮才智的进化主义, 生涯于不同环境(举例洪水vs缓流生境)的鱼类拍浮才智存在显着互异[29], 但同域漫步物种拍浮才智是否存在趋同适合尚无定论[30]。Raffini等[31]相关发现位于同域火山口湖的阿氏双冠丽鱼(Amphilophus astorquii)与札里双冠丽鱼(A. zaliosus)的急加快领路才智(Sprint swimming speed)无显贵互异, 但偏好稠密区的札里双冠丽鱼Ucrit高于喜底栖的阿氏双冠丽鱼、且Ucrit擢升与较高的氧需求关联, 异质栖息地中的氧供给互异可能在同域物种适合性分化中起雄壮作用; Myrick等[32]检会了不同温度下同域漫步物种臼齿鱼(Mylopharodon conocephalus)、细尾好意思拟鲴(Lavinia exilicauda)和丰润叶唇鱼(Ptychocheilus grandis)的拍浮才智, 发现它们具有左近的拍浮速率、且以肖似的神气对温度变化作出反馈, 这可能是由于相同环境选定压力形成的趋同进化所致; 而Yamada等[33]联紧缚尾则标明, 6 ℃时花羔红点鲑(Salvelinus malma)的拍浮才智强于同域漫步的白点鲑(Salvelinus leucomaenis)、但在12 ℃时拍浮才智无种间互异, 逢迎两者不同温度下上风地位发生转变的配景, 臆测拍浮才智受季节性温度变化的分化可能介导种间竞争末端, 从而影响二者共存; 此外, 对同域漫步且生态位重迭的食蚊鱼(Gambusia affinis)和唐鱼(Tanichthys albonubes)的相关发现, 永恒栖息于流动性更洪水环境中的唐鱼Ucat和Ucrit均高于生涯于水体流动较小生境中的食蚊鱼, 领导微生境选定分化可能驱动同域物种拍浮才智表型的种间互异[34]。本相关中, 总体上秦岭细鳞鲑Uburst高于拉氏鱥, 两物种Uburst互异显贵、不存在趋同适合。除遗传因素外, 咱们臆测生态习性不同可能是该同域物种爆发拍浮才智出现互异的主要原因。一方面, 秦岭细鳞鲑生殖与索饵洄游横暴、平庸需要穿越各式复杂激流生境、为洄游性鱼类, 而拉氏鱥则相对趋于假寓性、生涯范围变动不大; 另一方面, 秦岭细鳞鲑为肉食性鱼类且以拉氏鱥为主要饵料生物、更高的Uburst成心于擢升捕食到手率, 而拉氏鱥为杂食性鱼类、相较而言对爆发拍浮才智的依赖不彊。值得持重的是, 尽管秦岭细鳞鲑成体和亚成体Uburst均高于拉氏鱥(不管是其成体仍是亚成体), 在初度测定中秦岭细鳞鲑幼体与拉氏鱥成体和亚成体Uburst均无显贵互异, 其干系种间互作机制有待于进一步相关。
3.3 两种践诺鱼Uburst的种内变异(生活史阶段效应)物种生态性能平庸随生活史发育历程而动态变化[18, 35—36], 因此对溪流鱼类拍浮才智的评估应相等关心生活史阶段效应。Kopf等[37]测定了澳大利亚突吻麦鳕鲈(Maccullochella macquariensis)、虫纹麦鳕鲈(Maccullochella peelii)、金鲈(Macquaria ambigua)等6种土著淡水鱼类早期生活史阶段Ucrit和疲顿拍浮速率(Prolonged swimming speed), 发现Ucrit随幼鱼的滋长发育而加多, 且比较于体长、大小、年纪等因素, 发育阶段与拍浮才智的干系性更高; 肖似地, 高泉等[38]发现, 6月龄斑马鱼(Danio rerio)平时成体与30月龄朽迈期个体比较, 6月龄个体的Uburst、拍浮距离和握续时辰等均显贵高于朽迈期个体; 然而, Swanson等[39]发现, 生活史阶段效应酬越洋公鱼(Hypomesus transpacificus)Ucrit过头拍浮耐力均无显贵影响, 作家臆测该末端可能与践诺鱼体型较小、随生活史阶段变化不显着关联。本相关中, 生活史阶段效应酬秦岭细鳞鲑和拉氏鱥Uburst、rUburst均影响显贵; 两物种爆发拍浮才智种内变异方法左近, Uburst均随发育年纪加多而加多、呈现出“成体>亚成体>幼体”方法, rUburst均随发育年纪加多而减小、呈现出“成体 < 亚成体 < 幼体”方法。这种变化趋势与干系文件中对斑重唇鱼(Diptychus maculatus Steindachner)、青鱼(Mylopharyngodon piceus)和斯氏高原鳅(Triplophysa stoliczkae)等鱼类测定末详察一致[40]。追随发育历程, Uburst加多可能与机体能量储备、肌肉增长和末端加多等关联, 而rUburst减小则可能是机体名义积增大引起拍浮时水阻非线性加多等原因所致[41—42]。
综上, 本践诺研制了一种仿生态鱼类爆发拍浮才智测定安设, 通过对秦岭细鳞鲑过头同域物种拉氏鱥爆发拍浮才智的相关标明该步伐可靠性较高; 相较于拉氏鱥, 秦岭细鳞鲑爆发拍浮才智具有更强的规复力; 秦岭细鳞鲑和拉氏鱥爆发拍浮才智种内种间互异显贵校园春色 亚洲色图, 二者在种间水平上不存在趋同适合、在种内水平上均弘扬出显贵的生活史阶段效应。联紧缚尾领导, 将来鱼类拍浮才智关联的鱼谈瞎想或涉水工程评价中, 应详细接头生活史阶段效应和不同物种的生态习性过头种间互异。